
 Submit a Paper
Submit a Paper Propose a Special lssue
Propose a Special lssue Open Access
Open Access
REVIEW

Protein-mediated interactions in the dynamic regulation of acute inflammation
Department of Pediatric Critical Care Medicine, Vanderbilt University Medical Center, Nashville, TN, 37232-9075, USA
* Corresponding Author: RYAN STARK. Email:
(This article belongs to the Special Issue: Biochemical and Epigenetics Changes in Health and Disease)
BIOCELL 2023, 47(6), 1191-1198. https://doi.org/10.32604/biocell.2023.027838
Received 18 November 2022; Accepted 09 February 2023; Issue published 19 May 2023
 View Full Text
View Full Text Download PDF
Download PDFAbstract
Protein-mediated interactions are the fundamental mechanism through which cells regulate health and disease. These interactions require physical contact between proteins and their respective targets of interest. These targets include not only other proteins but also nucleic acids and other important molecules as well. These proteins are often involved in multibody complexes that work dynamically to regulate cellular health and function. Various techniques have been adapted to study these important interactions, such as affinity-based assays, mass spectrometry, and fluorescent detection. The application of these techniques has led to a greater understanding of how protein interactions are responsible for both the instigation and resolution of acute inflammatory diseases. These pursuits aim to provide opportunities to target specific protein interactions to alleviate acute inflammation.Keywords
The regulation of cellular processes by proteins forms the foundation of all cellular biology and pathobiology. Fixed or transient interactions of proteins with their respective targets of interest allow for broad, dynamic modifications of cell mechanisms for both health and disease (de Las Rivas and Fontanillo, 2010). Targets of these interactions include other proteins, RNA, DNA, carbohydrates, and lipids. These interactions, in turn, regulate enzymatic alterations, molecular transport, cell expression patterns, and global signal transduction. While proper protein-mediated interactions are necessary for cell homeostasis, the importance of these interactions is no less true in acute inflammatory conditions. Under these settings, the dynamic protein-mediated interactions provide the dual purpose of initiating an appropriate response to the inflammatory challenge but also a mechanism to temper the response when the threat is abated. These opposing responses ideally work in concert to control the response and restore homeostasis, with the survival of the host as the outcome. However, dysregulation of these responses can lead to enhanced or perturbed protein-mediated interactions, causing the host to succumb to the disease. This mini-review will provide broad strokes on the mechanisms of protein-mediated interactions with a variety of cellular components, how alterations in these interactions result from and produce disease in acute inflammatory conditions, and suggest potential therapeutic avenues to regulate these mechanisms to tamper disease states (Fig. 1).
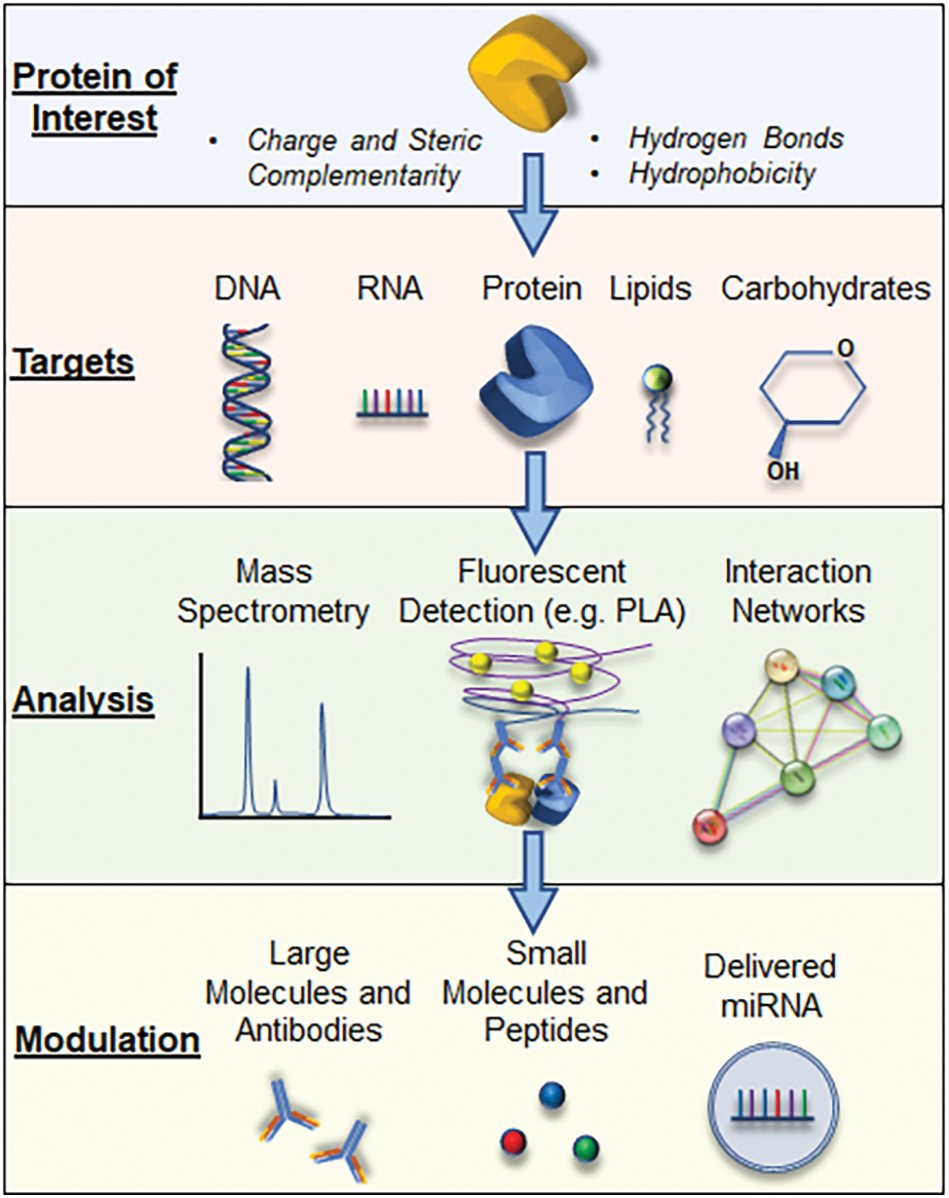
Figure 1: A schematic representation outlining how proteins of interest interact with possible binding partners and the potential therapeutic avenues that can be employed to modulate the known interactions.
Mechanisms of Protein-Mediated Interactions
In the broadest sense, protein-protein interactions form the mechanisms through which multi-protein complexes are created (Dinant et al., 2009). The interactions fundamentally depend on the amino acid sequences and the associated charge of the proteins forming the interaction. In some instances, gaps are formed in protein folding that allows for other molecules or proteins to fill. This process is called steric complementarity and forms the “lock-and-key” approach to protein binding (Blaney et al., 1982). Comparably, electrostatic charges of amino acid sequences form the basis for protein folding (Roca et al., 2007). When basic residues are in proximity to acidic residues, the electrostatic charge pulls the opposingly charged sequences together and can be used to form interactions between adjacent proteins. These interactions and steric relationships, create unique docking sites for protein-protein binding (Vakser, 2014). A vast number of these sequences, called conserved domains, have been cataloged and can be readily assessed (https://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml). In addition, both the hydrophobicity and shared bonding of hydrogen account for other mechanisms through which proteins can interact to modify one another (Jiang and Lai, 2002).
Protein-DNA and protein-RNA interactions
Similar to protein-protein interactions, it has been long recognized that proteins can form direct binding connections with nucleic acid structures (Blake and Oatley, 1977). Protein interactions with strands of various nucleic acid sequences are necessary for DNA-mediated production of proteins. Often these protein-mediated interactions modify various aspects of this process, from localization within the cell to sequence stability and editing (Re et al., 2014). But while the transcription of DNA to RNA and subsequent translation of RNA to amino acids is well understood, it is approximated that up to 80% of human DNA, and likewise RNA, do not code for proteins (Consortium, 2012). Instead, much of this genetic material has regulatory functions that include the modulation of the cellular environment through protein-mediated interactions (Elkon and Agami, 2017). In particular, non-coding RNA (ncRNA) are RNA sequences, either long (>200 bps) or short (<200 bps), that cannot code for a protein. Despite this, ncRNAs have been shown to regulate many aspects of cell biology, including through their regulation of proteins (Kazimierczyk et al., 2020). Understanding the role of these interactions is still in its infancy, but it has been suggested that the more complex a protein network is, the more likely it has interactions with ncRNAs, such as microRNAs (miRNAs), to alter the multi-protein complex function through dedicated ncRNA docking sites (Liang and Li, 2007).
Other direct protein interactions
Though protein-protein and protein-DNA/RNA interactions are important in signaling, proteins can interact with other cellular components, namely lipids, and carbohydrates. Like the mechanisms previously mentioned, protein-lipid or protein-carbohydrate interactions often depend on charge and hydrophobicity that allow for specific binding. In the case of carbohydrates, the many hydroxyl groups (−OH) present in carbohydrates allow for the potential of hydrogen bond crosslinking (Zhang et al., 2021). Likewise, electrostatic interactions and divalent cationic bridges also account for protein-carbohydrate interactions, similar to other protein-mediated mechanisms mentioned (Boraston et al., 2004). In a similar context, protein-lipid interactions are crucial for supporting cell membrane integrity. Binding sites found on proteins help form these interactions. In particular, pleckstrin homology domains on proteins have been well-established to bind phosphatidylinositol lipids, which aid in cytoskeleton rearrangements (Corey et al., 2020).
The capture of protein-mediated interactions can be challenging due to their dynamic and transient nature. Typically, for a protein-mediated interaction to be considered meaningful, the interaction should be considered intentional (i.e., not via Brownian motion) and with a specific biological purpose to induce a physiologic change (de Las Rivas and Fontanillo, 2010). Some more basic detection methods include affinity purification-mass spectrometry and co-immunoprecipitation, which identify protein-binding partners (Rao et al., 2014; Cozzolino et al., 2021). X-ray crystallography provides three-dimensional structure and visualization of interactions, making this technique exceptionally important in the lock-and-key design of drug targets. Other fluorescent microscopic techniques, such as proximity ligation assays (PLA) and förster resonance energy transfer (FRET), provide some spatial understanding of protein-mediated interactions (Ivanusic et al., 2014). Assessing the strength of these interactions can be performed with such techniques as atomic force microscopy (AFM) and biolayer interferometry (Whited and Park, 2014; Barrows and van Dyke, 2022). More recently, computational techniques have been developed to assess the complex and dynamic relationships that protein-mediated interactions have on cell function. These include computational models to predict possible interactions and large-scale databases to test known binding partners or interactions within biological networks (Ding and Kihara, 2018; Droit et al., 2005; Yang et al., 2020). For instance, the STRING Database (https://string-db.org/) allows for the analysis of both physical and functional interactions between multiple proteins of interest. Understanding the mechanisms of protein-mediated interactions, and the tools to assess them, have allowed for investigations into the relevance of these relationships during acute inflammation.
Protein-Mediated Interactions in Acute Inflammation
At the most basic level, protein-mediated interactions are the drivers of the cellular response to acute inflammatory challenges. Starting with either cytokines, damage-associated or pathogen-associated molecular patterns (DAMPs and PAMPs), interactions of these molecules with pattern recognition receptors (PRRs) start a signaling cascade. During an inflammatory challenge, this cascade is initiated and propagated by a series of protein-mediated processes that include protein-protein, protein-DNA/RNA, protein-carbohydrate, and protein-lipid interactions to drive the biological response (Schweppe et al., 2015). One example is lectins, which are non-enzymatic carbohydrate-binding proteins and important instigators and regulators of the host response to pathogens (Mason and Tarr, 2015). Indeed, C-type lectin receptors (CLRs) are one of the primary classes of PRRs that form a crucial first step to initiating an inflammatory response to pathogenic challenge (Geijtenbeek and Gringhuis, 2009). Another well-described class of PRRs is toll-like receptors (TLRs), which are homo or heterodimeric proteins that recognize foreign or endogenous proteins, lipids, or DNA/RNA to start the inflammatory response (Kawasaki and Kawai, 2014). Further protein-mediated interactions at the cellular surface include the previously mentioned protein-lipid interactions necessary for the movement of proteins on the cell surface and endocytosis-mediated signaling (Ewers and Helenius, 2011).
The protein-mediated signals that occur at the plasma membrane and beyond are typically the result of a series of assembling and dissembling multi-protein complexes that exert their effect on various parts of the cell. These multi-protein complexes aid in signal transduction through a variety of post-translational modifications to induce protein targets to carry out specific functions (Ramazi and Zahiri, 2021). Signal transduction through inflammatory cascades is also under the influence of either internal or external factors, such as non-protein coding RNA products. These products can, in turn, either enhance or abate the inflammatory response. The overall downstream protein-mediated processes related to this are far beyond the scope of this mini-review; however, the focus will be provided on two crucial aspects of inflammatory signaling: protein-RNA interactions in modulating the inflammatory response and protein-protein interactions in inflammation-mediated metabolism.
NcRNAs in pathogen-mediated inflammation
As mentioned previously, a majority of DNA does not code for a specific protein. Nevertheless, these non-coding regions have important regulatory functions in host defense (Li et al., 2021). In one recent study in neonates with sepsis, which is a severe immunological response to a pathogen, 89 long non-coding RNAs (lncRNAs) were found to be altered (Bu et al., 2020). One well-described lncRNA is metastasis-associated lung adenocarcinoma transcript 1 (MALAT1). Elevated circulating levels of MALAT1 have been shown in patients with sepsis and coronavirus disease (COVID)-19, and these levels correlate with the severity of the disease (Chen et al., 2020; Huang et al., 2022). MALAT1 is known to interact with several proteins involved in RNA processing and gene transcription; however, a study using a combination of RNA pull-down and mass spectrometry showed that MALAT1 has potentially up to 127 protein binding partners. In particular, MALAT1 binds to a protein called depleted in breast cancer 1 (DBC1) and in doing so, increases a protein called sirtuin 1 (SIRT1) which is a regulator of cellular metabolism (Chen et al., 2017). MALAT1 also functions as a miRNA sponge (Liu et al., 2019). The term “sponge” is used when direct interactions occur between a miRNA and another molecule, which inhibits the miRNA of interest from carrying out its regulatory function (Bak and Mikkelsen, 2014). A meta-analysis of samples from patients with sepsis demonstrated that 39 miRNAs were differentially regulated (Formosa et al., 2022). One of these, miR-125B, has been associated with MALAT1. Data has shown that as MALAT1 expression increased, miR-125B decreased in proportion, and this again correlated with disease severity (Le and Shi, 2022). In this sense, miRNA expression patterns have been suggested as biomarkers to delineate disease and the severity of illness (Dumache et al., 2015). But beyond expression patterns, miRNAs have functional roles mediated through direct interactions. Once produced in the nucleus, they can exert their effect locally, such as via histone modifications or RNA silencing with subsequent protein reduction. They can also be packaged into exosomes for more distal impact. TLR4, a well-studied PRR whose primary agonist is gram-negative lipopolysaccharide, produces an array of miRNAs when activated, but in turn, its signaling cascade is also modulated at various points by miRNAs (Szilagyi et al., 2019). Thus, it can be seen that non-protein coding RNA products are mechanisms through which the host fine-tunes the inflammatory response to a challenge via direct protein-RNA or RNA-RNA interactions.
Protein interactions in the regulation of inflammatory metabolism
The crux of a host response to combat a severe inflammatory challenge is to ensure that the energy needs of the cells are met (Lee and Huttemann, 2014). Under resting conditions, much of the energy produced as adenosine triphosphate (ATP) is through mitochondrial oxidative phosphorylation as opposed to glycolysis, the latter of which also generates ATP, though to a much lesser degree (Bonora et al., 2012). However, in times of stress, the phosphate group is quickly utilized, producing adenosine monophosphate and adenosine diphosphate, which must be readily restored to ATP. Though in the resting state, oxidative phosphorylation is the premier source of ATP generation, during an infectious challenge, the path of energy production pivots towards glycolysis (van Wyngene et al., 2018). This described “metabolic shift” is important in host defense and survival (Cheng et al., 2016). The processes that regulate this shift are highly dependent on protein-mediated interactions and disruptions can lead to an altered immunometabolic phenotype. Hypoxia-inducible factor 1 alpha (HIF-1α), a key regulator of metabolism during stress, is impacted by the capture of a protein called von Hippel–Lindau protein (VHL), leading to HIF-1α degradation (Weidemann and Johnson, 2008). Upregulation of HIF-1α occurs during sepsis, and its destruction by protein-protein interactions with VHL is thought to shift the cells toward fatty-acid metabolism and an anti-inflammation phenotype (Fitzpatrick, 2019). Similarly, SIRTs are another class of proteins that are thought to be important mediators of metabolism during times of acute stress. Of the seven subtypes of SIRTs known, SIRT1 is the best described in its link with metabolism (Chang and Guarente, 2014). SIRT1 has a vast array of proteins it interacts with through its deacetylation activity (McBurney et al., 2013). In endothelial cells, SIRT1 interacts with several key regulatory and homeostatic proteins (Stark et al., 2022). These interactions include not only those with metabolism-related functions, such as endothelial nitric oxide synthase and forkhead box O1, but also with structural proteins, such as vascular endothelial cadherin (VE-cadherin), putting SIRT1 protein networks at the crossroads of structure and function (Fig. 2). In total, alternations or disruptions in these protein-mediated interactions drive the clinical phenotypes seen and provide opportunities for potential therapeutic intervention.
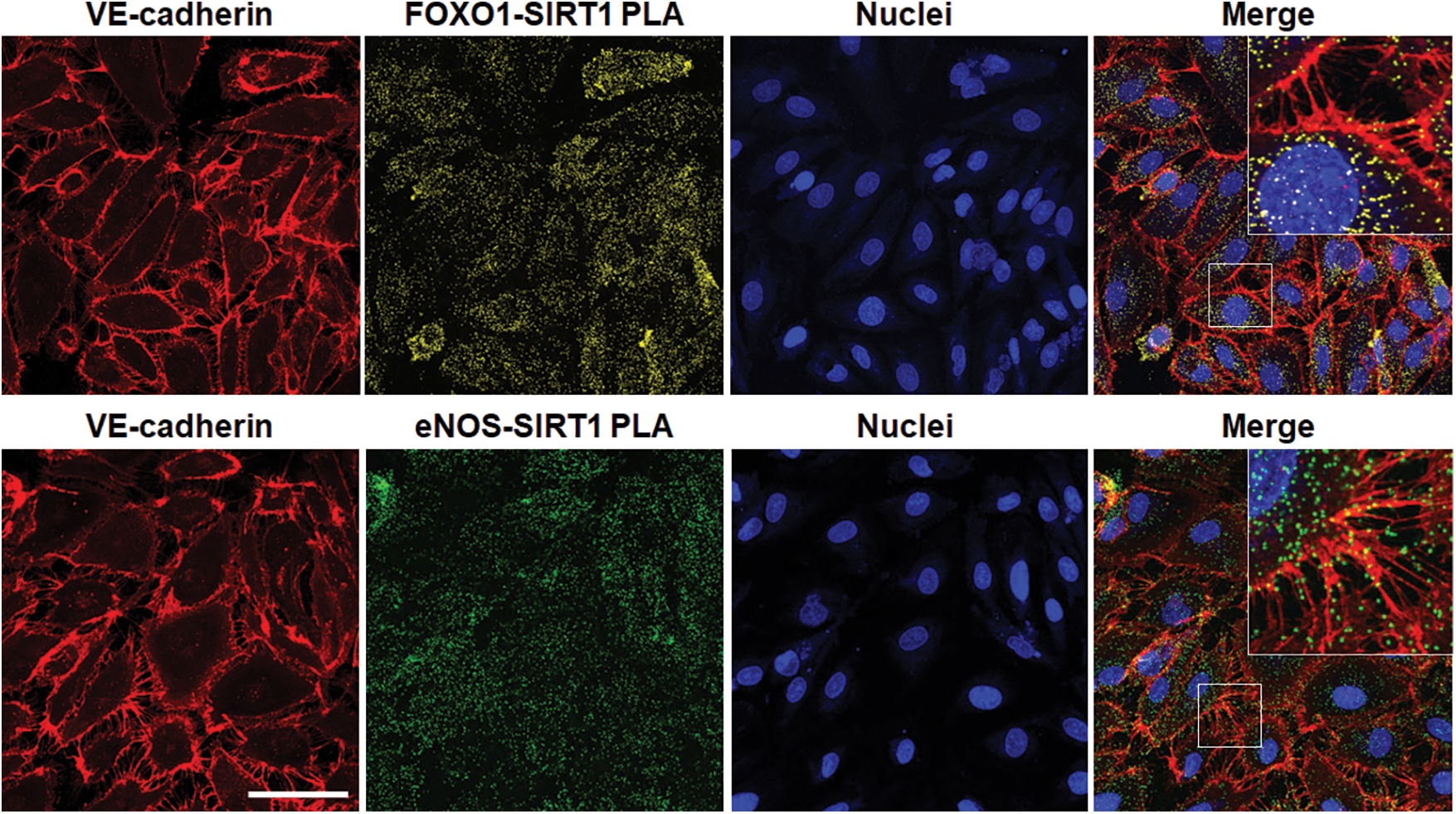
Figure 2: Demonstration of forkhead box 1 (FOXO1, top) or endothelial nitric oxide synthase (eNOS, bottom) and sirtuin 1 (SIRT1) interactions in human microvascular endothelial cells during acute challenge by gram-negative lipopolysaccharide (LPS). Immunofluorescence staining of endothelial VE-cadherin (red) and FOXO1-SIRT1 (yellow) or eNOS-SIRT1 (green) interactions by proximity ligation assay (PLA) with nuclei (blue). Merged images with inserts shown at the far-right demonstrate FOXO1-SIRT1 and eNOS-SIRT1 protein-protein interactions within the projections of VE-cadherin, necessary for maintaining tight gap junctions, between adjacent endothelial cells during pathogenic inflammatory challenge. Captured on a Zeiss LSM 880 inverted confocal microscope. Bar = 50 μm. Unpublished image courtesy of R. Stark derived from experiments outlined in Stark et al. (2022).
Methods of Targeting Protein Interactions
The importance of understanding protein-mediated interactions cannot be understated, especially in the context of drug discovery to mitigate disease. Potentiating or inhibiting key interactions during a disease process provides the possibility to hasten or restore homeostasis. As mentioned previously, the foundation of most protein-mediated interactions involves a lock-and-key interaction. Interactions can be directly targeted through orthosteric mechanisms or via indirect targeting through allosteric mechanisms (Nussinov and Tsai, 2012). Lu et al. (2020) have already provided an excellent and thorough review of this subject matter, so this discussion will focus on key highlights and generalized methods of targeting protein interactions, especially within the context of acute inflammation.
The use of large-molecule biologics to regulate inflammation has been well-established for chronic inflammatory conditions. In these settings, the biologics act as scavengers of pro-inflammatory soluble molecules circulating in the blood or as receptor antagonists (Rider et al., 2016). Many of these biological substances come in the form of monoclonal antibodies. Unfortunately, due to their long half-life and the necessity of proper cytokine signaling in acute inflammatory conditions, the application of these therapeutics comes with a significant risk of secondary infections or reactivations (Lang et al., 2012). However, large molecules with shorter half-lives may be of some utility. This includes drugs such as anakinra, an interleukin-1 receptor antagonist that has an elimination half-life of 6 hours and has been long used in the treatment of autoimmune diseases (Bedaiwi et al., 2021). Given its relatively good safety profile, it has been used in acute inflammatory conditions. Though global results have not been promising, selectivity in patient selection based on immunological profiling may provide more encouraging results (Fisher et al., 1994; Shakoory et al., 2016).
Small molecule inhibitors and peptides
The utility of designing drugs to interfere with protein-mediated interactions was never made clearer than during the pandemic associated with COVID-19. This resurgence of interest led to the application of small molecule compounds designed to interfere with the interactions of coronavirus with the host. Remdesivir, a nucleoside analog that interferes with the RNA-dependent RNA polymerase complex used in viral replication, was quickly approved for use in COVID-19 (Kokic et al., 2021). The small molecule Janus kinase 1/2 inhibitor, known as baricitinib, was also found to be effective with increased efficacy when combined with remdesivir in patients with COVID-19 (Kalil et al., 2021). An additional trial (NCT04311697) using synthetic vasoactive intestinal peptide (aviptadil) in the treatment of COVID-19 is currently awaiting results, overall demonstrating the feasibility and efficacy of these therapeutics in acute inflammatory diseases. Beyond this, the discovery of future small molecule compounds is likely to accelerate with the development of high-throughput compound screening libraries. For instance, utilizing such a library revealed that niclosamide, approved as an anti-helminthic drug, had broad anti-inflammatory properties via inflammasome inhibition that worked against coronavirus, as well as several different TLR ligands (de Almeida et al., 2022). Similarly, cell-penetrating peptides (CPPs), along with naturally occurring antimicrobial peptides, have been suggested to have therapeutic potential in modulating protein-mediated interactions. Several compounds have been shown to affect intracellular pathogens as well as regulate infection-mediated endothelial barrier dysfunction (Buccini et al., 2020; Koch and Stark, 2022). However, despite the pre-clinical promise of CPPs, none have been approved for clinical use thus far (Xie et al., 2020).
Manipulation of protein production and signal transduction by miRNAs has been previously discussed. Given the wide impact miRNAs can have on proteins, it makes them an attractive potential therapeutic avenue for targeting cellular processes (Rupaimoole and Slack, 2017). Data thus far regarding the therapeutic potential of miRNAs in the treatment of disease have been limited. Several clinical trials utilizing miRNAs have been performed targeting cancers or chronic viral infections (Janssen et al., 2013; Hong et al., 2020). In addition, there are several ongoing clinical trials targeting miRNAs to improve outcomes in ulcerative colitis (NCT04023396) or myocardial infarction (NCT05350969). While currently there are no miRNA therapeutics aimed at regulating acute, acquired inflammatory diseases, such as sepsis or COVID-19, the concept has gained some traction (Giza et al., 2016). Regarding COVID-19, there is currently an ongoing clinical trial examining the use of exosomes containing miRNA to treat COVID-19-mediated inflammation (NCT05216562). Though the therapeutic application of miRNAs to successfully treat acute inflammatory diseases remains to be seen, they nevertheless continue to be a topic of significant investment.
Great strides have been made in recent decades that have begun to pull back the veil of complexity that is the host’s defense to acute inflammatory challenge. With the advent of more sophisticated techniques, coupled with enhanced computing tools and machine learning, we now know more about the interconnected and dynamic proteome involved in modulating a host’s response. This knowledge has been further expanded by understanding new facets of the role of the genome in producing non-coding RNA, which regulate protein structure and function, as well as the importance of multi-protein complexes forming the appropriate metabolic response to an inflammatory challenge. As these protein-mediated interactions are further elucidated, high-throughput screening techniques that repurpose old drugs, or allow for the creation of new ones, will hopefully allow for the correction of aberrant protein interactions that instigate and perpetuate acute inflammatory conditions. The development of such compounds will no doubt usher in a new era of therapeutics that will provide improved outcomes for those patients suffering from diseases associated with acute inflammation.
Funding Statement: This work was supported by a grant from the National Institutes of Health [R35 GM138191 to RS].
Author Contributions:: The author confirms sole responsibility for the following: study conception and design, data collection, analysis and interpretation of results, and manuscript preparation.
Conflicts of Interest: The author declares that they have no conflicts of interest to report regarding the present study.
References
Bak RO, Mikkelsen JG (2014). miRNA sponges: Soaking up miRNAs for regulation of gene expression. WIREs RNA 5: 317–333. https://doi.org/10.1002/wrna.1213 [Google Scholar] [PubMed] [CrossRef]
Barrows JK, Van Dyke MW (2022). Biolayer interferometry for DNA-protein interactions. PLoS One 17: e0263322. https://doi.org/10.1371/journal.pone.0263322 [Google Scholar] [PubMed] [CrossRef]
Bedaiwi MK, Almaghlouth I, Omair MA (2021). Effectiveness and adverse effects of anakinra in treatment of rheumatoid arthritis: A systematic review. European Review for Medical and Pharmacological Sciences 25: 7833–7839. https://doi.org/10.26355/eurrev_202112_27630 [Google Scholar] [PubMed] [CrossRef]
Blake CC, Oatley SJ (1977). Protein-DNA and protein-hormone interactions in prealbumin: A model of the thyroid hormone nuclear receptor? Nature 268: 115–120. https://doi.org/10.1038/268115a0 [Google Scholar] [PubMed] [CrossRef]
Blaney JM, Jorgensen EC, Connolly ML, Ferrin TE, Langridge R, Oatley SJ, Burridge JM, Blake CC (1982). Computer graphics in drug design: molecular modeling of thyroid hormone-prealbumin interactions. Journal of Medicinal Chemistry 25: 785–790. https://doi.org/10.1021/jm00349a004 [Google Scholar] [PubMed] [CrossRef]
Bonora M, Patergnani S, Rimessi A, de Marchi E, Suski JM et al. (2012). ATP synthesis and storage. Purinergic Signal 8: 343–357. https://doi.org/10.1007/s11302-012-9305-8 [Google Scholar] [PubMed] [CrossRef]
Boraston AB, Bolam DN, Gilbert HJ, Davies GJ (2004). Carbohydrate-binding modules: Fine-tuning polysaccharide recognition. Biochemical Journal 382: 769–781. https://doi.org/10.1042/BJ20040892 [Google Scholar] [PubMed] [CrossRef]
Bu L, Wang ZW, Hu SQ, Zhao WJ, Geng XJ et al. (2020). Identification of key mRNAs and lncRNAs in neonatal sepsis by gene expression profiling. Computational and Mathematical Methods in Medicine 2020: 8741739. https://doi.org/10.1155/2020/8741739 [Google Scholar] [PubMed] [CrossRef]
Buccini DF, Cardoso MH, Franco OL (2020). Antimicrobial peptides and cell-penetrating peptides for treating intracellular bacterial infections. Frontiers in Cellular and Infection Microbiology 10: 612931. https://doi.org/10.3389/fcimb.2020.612931 [Google Scholar] [PubMed] [CrossRef]
Chang HC, Guarente L (2014). SIRT1 and other sirtuins in metabolism. Trends in Endocrinology & Metabolism 25: 138–145. https://doi.org/10.1016/j.tem.2013.12.001 [Google Scholar] [PubMed] [CrossRef]
Chen J, He Y, Zhou L, Deng Y, Si L (2020). Long noncoding RNA MALAT1 serves as an independent predictive biomarker for the diagnosis, severity and prognosis of patients with sepsis. Molecular Medicine Reports 21: 1365–1373. https://doi.org/10.3892/mmr.2020.10923 [Google Scholar] [PubMed] [CrossRef]
Chen R, Liu Y, Zhuang H, Yang B, Hei K et al. (2017). Quantitative proteomics reveals that long non-coding RNA MALAT1 interacts with DBC1 to regulate p53 acetylation. Nucleic Acids Research 45: 9947–9959. https://doi.org/10.1093/nar/gkx600 [Google Scholar] [PubMed] [CrossRef]
Cheng SC, Scicluna BP, Arts RJ, Gresnigt MS, Lachmandas E et al. (2016). Broad defects in the energy metabolism of leukocytes underlie immunoparalysis in sepsis. Nature Immunology 17: 406–413. https://doi.org/10.1038/ni.3398 [Google Scholar] [PubMed] [CrossRef]
Consortium EP (2012). An integrated encyclopedia of DNA elements in the human genome. Nature 489: 57–74. https://doi.org/10.1038/nature11247 [Google Scholar] [PubMed] [CrossRef]
Corey RA, Stansfeld PJ, Sansom MSP (2020). The energetics of protein-lipid interactions as viewed by molecular simulations. Biochemical Society Transactions 48: 25–37. https://doi.org/10.1042/BST20190149 [Google Scholar] [PubMed] [CrossRef]
Cozzolino F, Iacobucci I, Monaco V, Monti M (2021). Protein-DNA/RNA Interactions: An overview of investigation methods in the -omics era. Journal of Proteome Research 20: 3018–3030. https://doi.org/10.1021/acs.jproteome.1c00074 [Google Scholar] [PubMed] [CrossRef]
de Almeida L, da Silva ALN, Rodrigues TS, Oliveira S, Ishimoto AY et al. (2022). Identification of immunomodulatory drugs that inhibit multiple inflammasomes and impair SARS-CoV-2 infection. Science Advances 8: eabo5400. https://doi.org/10.1126/sciadv.abo5400 [Google Scholar] [PubMed] [CrossRef]
De Las Rivas J, Fontanillo C (2010). Protein-protein interactions essentials: Key concepts to building and analyzing interactome networks. PLoS Computational Biology 6: e1000807. https://doi.org/10.1371/journal.pcbi.1000807 [Google Scholar] [PubMed] [CrossRef]
Dinant C, Luijsterburg MS, Hofer T, von Bornstaedt G, Vermeulen W, Houtsmuller AB, van Driel R (2009). Assembly of multiprotein complexes that control genome function. Journal of Cell Biology 185: 21–26. https://doi.org/10.1083/jcb.200811080 [Google Scholar] [PubMed] [CrossRef]
Ding Z, Kihara D (2018). Computational methods for predicting protein-protein interactions using various protein features. Current Protocols in Protein Science 93: e62. https://doi.org/10.1002/cpps.62 [Google Scholar] [PubMed] [CrossRef]
Droit A, Poirier GG, Hunter JM (2005). Experimental and bioinformatic approaches for interrogating protein-protein interactions to determine protein function. Journal of Molecular Endocrinology 34: 263–280. https://doi.org/10.1677/jme.1.01693 [Google Scholar] [PubMed] [CrossRef]
Dumache R, Rogobete AF, Bedreag OH, Sarandan M, Cradigati AC, Papurica M, Dumbuleu CM, Nartita R, Sandesc D (2015). Use of miRNAs as biomarkers in sepsis. Analytical Cellular Pathology 2015: 186716. https://doi.org/10.1155/2015/186716 [Google Scholar] [PubMed] [CrossRef]
Elkon R, Agami R (2017). Characterization of noncoding regulatory DNA in the human genome. Nature Biotechnology 35: 732–746. https://doi.org/10.1038/nbt.3863 [Google Scholar] [PubMed] [CrossRef]
Ewers H, Helenius A (2011). Lipid-mediated endocytosis. Cold Spring Harbor Perspectives in Biology 3: a004721. https://doi.org/10.1101/cshperspect.a004721 [Google Scholar] [PubMed] [CrossRef]
Fisher Jr CJ, Dhainaut JF, Opal SM, Pribble JP, Balk RA et al. (1994). Recombinant human interleukin 1 receptor antagonist in the treatment of patients with sepsis syndrome. Results from a randomized, double-blind, placebo-controlled trial. Phase III rhIL-1ra Sepsis Syndrome Study Group. JAMA 271: 1836–1843. https://doi.org/10.1001/jama.1994.03510470040032 [Google Scholar] [CrossRef]
Fitzpatrick SF (2019). Immunometabolism and sepsis: A role for HIF? Frontiers in Molecular Biosciences 6: 85. https://doi.org/10.3389/fmolb.2019.00085 [Google Scholar] [PubMed] [CrossRef]
Formosa A, Turgeon P, Dos Santos CC (2022). Role of miRNA dysregulation in sepsis. Molecular Medicine 28: 99. https://doi.org/10.1186/s10020-022-00527-z [Google Scholar] [PubMed] [CrossRef]
Geijtenbeek TB, Gringhuis SI (2009). Signalling through C-type lectin receptors: Shaping immune responses. Nature Reviews: Immunology 9: 465–479. https://doi.org/10.1038/nri2569 [Google Scholar] [CrossRef]
Giza DE, Fuentes-Mattei E, Bullock MD, Tudor S, Goblirsch MJ, Fabbri M, Lupu F, Yeung SJ, Vasilescu C, Calin GA (2016). Cellular and viral microRNAs in sepsis: Mechanisms of action and clinical applications. Cell Death and Differentiation 23: 1906–1918. https://doi.org/10.1038/cdd.2016.94 [Google Scholar] [PubMed] [CrossRef]
Hong DS, Kang YK, Borad M, Sachdev J, Ejadi S et al. (2020). Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. British Journal of Cancer 122: 1630–1637. https://doi.org/10.1038/s41416-020-0802-1 [Google Scholar] [PubMed] [CrossRef]
Huang K, Wang C, Vagts C, Raguveer V, Finn PW, Perkins DL (2022). Long non-coding RNAs (lncRNAs) NEAT1 and MALAT1 are differentially expressed in severe COVID-19 patients: An integrated single-cell analysis. PLoS One 17: e0261242. https://doi.org/10.1371/journal.pone.0261242 [Google Scholar] [CrossRef]
Ivanusic D, Eschricht M, Denner J (2014). Investigation of membrane protein-protein interactions using correlative FRET-PLA. Biotechniques 57: 188–198. https://doi.org/10.2144/000114215 [Google Scholar] [PubMed] [CrossRef]
Janssen HL, Reesink HW, Lawitz EJ, Zeuzem S, Rodriguez-Torres M et al. (2013). Treatment of HCV infection by targeting microRNA. New England Journal of Medicine 368: 1685–1694. https://doi.org/10.1056/NEJMoa1209026 [Google Scholar] [PubMed] [CrossRef]
Jiang L, Lai L (2002). CH..O hydrogen bonds at protein-protein interfaces. Journal of Biological Chemistry 277: 37732–37740. https://doi.org/10.1074/jbc.M204514200 [Google Scholar] [PubMed] [CrossRef]
Kalil AC, Patterson TF, Mehta AK, Tomashek KM, Wolfe CR et al. (2021). Baricitinib plus remdesivir for hospitalized adults with COVID-19. New England Journal of Medicine 384: 795–807. https://doi.org/10.1056/NEJMoa2031994 [Google Scholar] [PubMed] [CrossRef]
Kawasaki T, Kawai T (2014). Toll-like receptor signaling pathways. Frontiers in Immunology 5: 461. https://doi.org/10.3389/fimmu.2014.00461 [Google Scholar] [PubMed] [CrossRef]
Kazimierczyk M, Kasprowicz MK, Kasprzyk ME, Wrzesinski J (2020). Human long noncoding RNA interactome: Detection, characterization and function. International Journal of Molecular Sciences 21: 1027. https://doi.org/10.3390/ijms21031027 [Google Scholar] [PubMed] [CrossRef]
Koch SR, Stark RJ (2022). Cell penetrating peptides coupled to an endothelial nitric oxide synthase sequence alter endothelial permeability. Tissue Barriers 10: 2017226. https://doi.org/10.1080/21688370.2021.2017226 [Google Scholar] [PubMed] [CrossRef]
Kokic G, Hillen HS, Tegunov D, Dienemann C, Seitz F, Schmitzova J, Farnung L, Siewert A, Hobartner C, Cramer P (2021). Mechanism of SARS-CoV-2 polymerase stalling by remdesivir. Nature Communications 12: 279. https://doi.org/10.1038/s41467-020-20542-0 [Google Scholar] [PubMed] [CrossRef]
Lang VR, Englbrecht M, Rech J, Nusslein H, Manger K et al. (2012). Risk of infections in rheumatoid arthritis patients treated with tocilizumab. Rheumatology 51: 852–857. https://doi.org/10.1093/rheumatology/ker223 [Google Scholar] [PubMed] [CrossRef]
Le Y, Shi Y (2022). MALAT1 regulates PCT expression in sepsis patients through the miR-125b/STAT3 axis. Journal of Clinical Laboratory Analysis 36: e24428. https://doi.org/10.1002/jcla.24428 [Google Scholar] [PubMed] [CrossRef]
Lee I, Huttemann M (2014). Energy crisis: The role of oxidative phosphorylation in acute inflammation and sepsis. Biochimica et Biophysica Acta 1842: 1579–1586. https://doi.org/10.1016/j.bbadis.2014.05.031 [Google Scholar] [PubMed] [CrossRef]
Li J, Zhang Y, Zhang D, Li Y (2021). The role of long non-coding RNAs in sepsis-induced cardiac dysfunction. Frontiers in Cardiovascular Medicine 8: 684348. https://doi.org/10.3389/fcvm.2021.684348 [Google Scholar] [PubMed] [CrossRef]
Liang H, Li WH (2007). MicroRNA regulation of human protein protein interaction network. RNA 13: 1402–1408. https://doi.org/10.1261/rna.634607 [Google Scholar] [PubMed] [CrossRef]
Liu S, Qiu J, He G, Liang Y, Wang L, Liu C, Pan H (2019). LncRNA MALAT1 acts as a miR-125a-3p sponge to regulate FOXM1 expression and promote hepatocellular carcinoma progression. Journal of Cancer 10: 6649–6659. https://doi.org/10.7150/jca.29213 [Google Scholar] [PubMed] [CrossRef]
Lu H, Zhou Q, He J, Jiang Z, Peng C, Tong R, Shi J (2020). Recent advances in the development of protein-protein interactions modulators: Mechanisms and clinical trials. Signal Transduction and Targeted Therapy 5: 213. https://doi.org/10.1038/s41392-020-00315-3 [Google Scholar] [PubMed] [CrossRef]
Mason CP, Tarr AW (2015). Human lectins and their roles in viral infections. Molecules 20: 2229–2271. https://doi.org/10.3390/molecules20022229 [Google Scholar] [PubMed] [CrossRef]
McBurney MW, Clark-Knowles KV, Caron AZ, Gray DA (2013). SIRT1 is a highly networked protein that mediates the adaptation to chronic physiological stress. Genes & Cancer 4: 125–134. https://doi.org/10.1177/1947601912474893 [Google Scholar] [PubMed] [CrossRef]
Nussinov R, Tsai CJ (2012). The different ways through which specificity works in orthosteric and allosteric drugs. Current Pharmaceutical Design 18: 1311–1316. https://doi.org/10.2174/138161212799436377 [Google Scholar] [PubMed] [CrossRef]
Ramazi S, Zahiri J (2021). Posttranslational modifications in proteins: Resources, tools and prediction methods. Database 2021. https://doi.org/10.1093/database/baab012 [Google Scholar] [PubMed] [CrossRef]
Rao VS, Srinivas K, Sujini GN, Kumar GN (2014). Protein-protein interaction detection: Methods and analysis. International Journal of Proteomics 2014: 147648. https://doi.org/10.1155/2014/147648 [Google Scholar] [PubMed] [CrossRef]
Re A, Joshi T, Kulberkyte E, Morris Q, Workman CT (2014). RNA-protein interactions: An overview. In: Gorodkin J, Ruzzo WL (eds.RNA Sequence, Structure, and Function: Computational and Bioinformatic Methods. Totowa, NJ: Humana Press. [Google Scholar]
Rider P, Carmi Y, Cohen I (2016). Biologics for targeting inflammatory cytokines, clinical uses, and limitations. International Journal of Cell Biology 2016: 9259646. https://doi.org/10.1155/2016/9259646 [Google Scholar] [PubMed] [CrossRef]
Roca M, Messer B, Warshel A (2007). Electrostatic contributions to protein stability and folding energy. FEBS Letters 581: 2065–2071. https://doi.org/10.1016/j.febslet.2007.04.025 [Google Scholar] [PubMed] [CrossRef]
Rupaimoole R, Slack FJ (2017). MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nature Reviews Drug Discovery 16: 203–222. https://doi.org/10.1038/nrd.2016.246 [Google Scholar] [PubMed] [CrossRef]
Schweppe DK, Harding C, Chavez JD, Wu X, Ramage E, Singh PK, Manoil C, Bruce JE (2015). Host-microbe protein interactions during bacterial infection. Chemistry and Biology 22: 1521–1530. https://doi.org/10.1016/j.chembiol.2015.09.015 [Google Scholar] [PubMed] [CrossRef]
Shakoory B, Carcillo JA, Chatham WW, Amdur RL, Zhao H, Dinarello CA, Cron RQ, Opal SM (2016). Interleukin-1 receptor blockade is associated with reduced mortality in sepsis patients with features of macrophage activation syndrome: Reanalysis of a prior phase III trial. Critical Care Medicine 44: 275–281. https://doi.org/10.1097/CCM.0000000000001402 [Google Scholar] [PubMed] [CrossRef]
Stark RJ, Koch SR, Stothers CL, Pourquoi A, Lamb CK, Miller MR, Choi H (2022). Loss of Sirtuin 1 (SIRT1) potentiates endothelial dysfunction via impaired glycolysis during infectious challenge. Clinical and Translational Medicine 12: e1054. https://doi.org/10.1002/ctm2.1054 [Google Scholar] [PubMed] [CrossRef]
Szilagyi B, Fejes Z, Pocsi M, Kappelmayer J, Nagy Jr B (2019). Role of sepsis modulated circulating microRNAs. Electronic Journal of the International Federation of Clinical Chemistry and Laboratory Medicine 30: 128–145. Retrieved from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6599195/ [Google Scholar] [PubMed]
Vakser IA (2014). Protein-protein docking: From interaction to interactome. Biophysical Journal 107: 1785–1793. https://doi.org/10.1016/j.bpj.2014.08.033 [Google Scholar] [PubMed] [CrossRef]
Van Wyngene L, Vandewalle J, Libert C (2018). Reprogramming of basic metabolic pathways in microbial sepsis: Therapeutic targets at last? EMBO Molecular Medicine 10: e8712. https://doi.org/10.15252/emmm.201708712 [Google Scholar] [PubMed] [CrossRef]
Weidemann A, Johnson RS (2008). Biology of HIF-1alpha. Cell Death and Differentiation 15: 621–627. https://doi.org/10.1038/cdd.2008.12 [Google Scholar] [PubMed] [CrossRef]
Whited AM, Park PS (2014). Atomic force microscopy: A multifaceted tool to study membrane proteins and their interactions with ligands. Biochimica et Biophysica Acta 1838: 56–68. https://doi.org/10.1016/j.bbamem.2013.04.011 [Google Scholar] [PubMed] [CrossRef]
Xie J, Bi Y, Zhang H, Dong S, Teng L, Lee RJ, Yang Z (2020). Cell-penetrating peptides in diagnosis and treatment of human diseases: From preclinical research to clinical application. Frontiers in Pharmacology 11: 697. https://doi.org/10.3389/fphar.2020.00697 [Google Scholar] [PubMed] [CrossRef]
Yang M, Derbyshire MK, Yamashita RA, Marchler-Bauer A (2020). NCBI’s conserved domain database and tools for protein domain analysis. Current Protocols in Bioinformatics 69: e90. https://doi.org/10.1002/cpbi.90 [Google Scholar] [PubMed] [CrossRef]
Zhang S, Chen KY, Zou X (2021). Carbohydrate-protein interactions: Advances and challenges. Communications in Information and Systems 21: 147–163. https://doi.org/10.4310/CIS.2021.v21.n1.a7 [Google Scholar] [PubMed] [CrossRef]
Cite This Article
 Copyright © 2023 The Author(s). Published by Tech Science Press.
Copyright © 2023 The Author(s). Published by Tech Science Press.This work is licensed under a Creative Commons Attribution 4.0 International License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
 Downloads
Downloads
 Citation Tools
Citation Tools